











![]()
![]()
![]()
![]()
Заболевания крови
Использование мумие в стоматологии
Заболевания органов дыхания
Заболевания желудочно-кишечного тракта. Колиты
Язвенная болезнь желудка и двенадцатиперстной кишки
Периферическая нервная система. Экземы.
Заболевания печени
Заболевания нервной системы
Невриты лицевого нерва
Противовоспалительное действие мумие
Переломы
Ожоги
Трофические язвы
Побочные действия мумие
Антимикробное действие мумие
Гнойно-воспалительные процессы
Действие мумие на имунную систему
Влияние мумие на печень
«Весь мир беременеет одними и теми же идеями» и это естественно, так как принципы научного поиска одинаковы. Нередко к концу работы автор вдруг обнаруживает что аналогичный результат уже опубликован. Конечно, авторское самолюбие при этом страдает — не я первый. Однако, и второе, и третье доказательство какого-либо факта также необходимы. В науке фактом признается лишь то, что подтверждено другими исследователями...
Многие наши исследования подтвердили данные других авторов и послужили доказательством того, что Саянское мумие не отличается по своему действию от других «сортов», но экспериментальные работы, посвященные функции печени, даже сегодня, через 20 лет после того, как они были проведены, не утратили своей оригинальности.
Печень является одним из важнейших органов. Она не только связывает и нейтрализует большинство токсических продуктов, участвует в обмене белков, витаминов, гормонов, жиров, углеводов. Печень является как-бы главной фабрикой-"кухней»- организма. Все пищевые продукты, поступая в кровь, расщепляются до простейших три- и дикарбоновых кислот и образуют общий резерв организма — так называемый обменный пул (Кребс и др.). По мере необходимости из этих составных частей в организме вновь синтезируются белки, жиры и углеводы и осуществляется это в основном печенью. Поэтому выяснить влияние мумие на печень представлялось особенно важным.
Влиянию на функцию печени неспецифических биогенных стимуляторов пентоксила и метацила посвящено значительноe количество работ. Так, Г. Л. Билич (1966) применил пентоксил у детей с различными гнойными заболеваниями до и после операции и установил, что функция печени, нарушенная у больных пропорционально тяжести основного заболевания тяжести операционной травмы и наркоза, обычно восстанавливается на 7—9 день, а при лечении пентоксилом — на 3-4 день. В. Л. Арзяева, вызывая у крыс и мышей токсический гепатит, обнаружила, что лечение пентоксилом и метацилом достоверно повышает выживаемость животных, а функциональные и морфологические изменения в печени при этом значительно менее выражены. А. Г. Сатарова (1966), применяя метацил, сократила сроки лечения больных эпиде-I уическим гепатитом на 1/3. В. И. Витлин (1970) установил, что у животных с перевязанным общим желчным протоком (механическая желтуха) дистрофические изменения в печени при лечении метацилом развиваются значительно позже. Д. В. Усов с соавторами (1966) и М. В. Нацюк (1970) пока-I зал и благотворное действие пиримидинов при травме печени в эксперименте и при лечении больных с хроническими гепатитами и холециститами.
Большая работа проведена И. М. Солопаевой с соавторами(1966, 1969). Вводя крысам после резекции 60% ткани печени по 200 мг/кг пентоксила, они обнаружили усиление ми-готической активности клеток, усиление утилизации печенью резервных белков крови и т. д.
Мы изучили влияние мумие и пентоксила на морфологию инекоторые функции печени в опытах на 296 белых крысах. В первой серии опытов было 200 интактных животных. 30 из них получали в течение 10 дней пентоксил по 100 мг/кг, 125—различные дозы мумие и 45 — получали физиологический раствор и являлись контрольными.
Во второй серии опытов 60 крысам производили резекцию левой доли печени (33—35% массы органа); 22 из них были контрольными, а по 19 животным получали мумие или пентоксил по 100 мг/кг.
В третьей серии опытов у 36 крыс вызывали токсический гепатит путем десятидневного введения им НАК (нитрил-акриловая кислота), являющейся сильным гепатотропным ядом. Лечение также проводили мумие и пентоксилом. В опытных и контрольной группе было по 12 животных. Препараты, как и в других случаях, вводили через зонд в желудок на протяжении всего опыта. Начинали введение за 3 дня до затравки.
Всем животным проводились гистоморфологические исследования ткани печени, определяли ее относительный вес, содержание в ней белка, жиров, гликогена и некоторых микроэлементов. Определяли количество нуклеиновых кислот, уровень гликолиза и дыхание митохондрий. Кроме того, определяли общий белок и белковые фракции, Р-липопротеиды и . трансаминазы сыворотки и кортикостерон плазмы.
Как видно из таблицы 16, у интактных крыс, получавших на протяжении 10 дней пентоксил, увеличивается в печени содержание общего азота и снижается на 16—18% уровень остаточного азота. В результате содержание белка в печени возрастает, а коэффициент протеолиза уменьшается на 1/4. Эти данные совпадают с имеющимися в литературе. Соответственно, наблюдается и достоверное увеличение относительной массы печени.
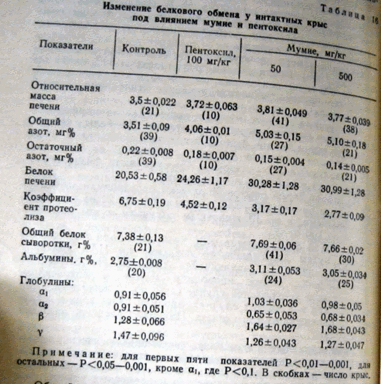
Таблица 16
Изменение белкового обмена у интактных крыс под влиянием мумие к пентоксила
Примечание: для первых пяти показателей Р<0,01—0,001, для остальных — Р<0,05—0,001, кроме a1, где Р<0,1. В скобках — число крыс.
Общий белок сыворотки, белковые фракции и бетта-липопротеиды изменяются несущественно. AS- и AL-трансаминазы снижаются на 37%. Содержание нуклеиновых кислот практически не изменяется (табл. 17).
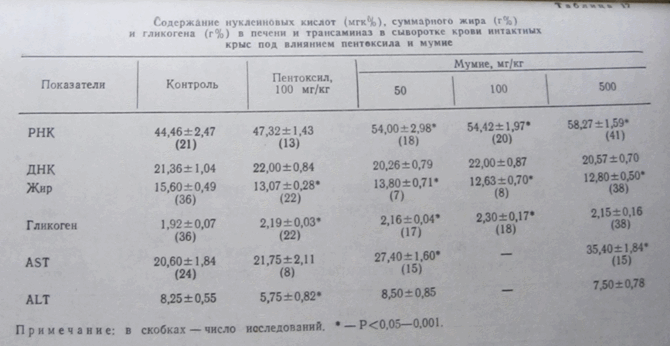
Под влиянием мумие происходят аналогичные и несколько более выраженные изменения, причем увеличение дозы мумие в 10 раз фактически не приводит к увеличению этих показателей. Полученные данные убедительно свидетельством, что мумие, так же как пиримидиновые производные, оказывает на печень выраженное анаболическое и антикаталическое действие.
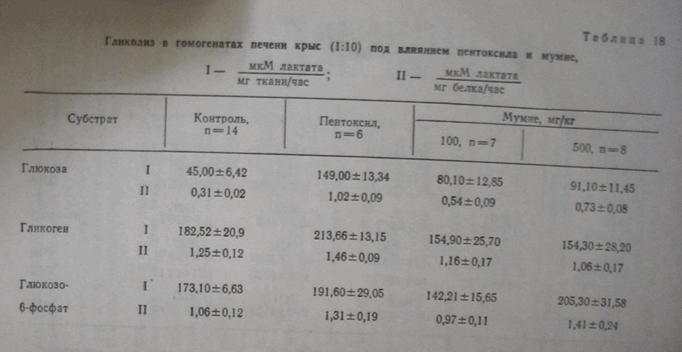
Одновременно отмечается и повышение белоксинтезирующей функции печени: в сыворотке повышается уровень общего белка, в белковых фракциях увеличивается содержание альбуминов и b-глобулинов, при снижении а2-глобулинов нa 25% (рис. 12). Остается непонятным снижение уровня
Рис. 12. Изменения общего белка, и белковых фракций сыворотки крови здоровых крыс под влиянием мумие.
Y-глобулинов в сыворотке крови, поскольку гистофункциональные исследования печени свидетельствуют о высокой активности купферовских клеток печени, а, как уже было описано
превышении липолиза над липогенезом и также подтверждается гистофункциональными исследованиями.
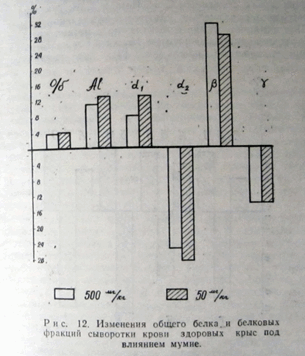
Мумие и пентоксил активно изменяют гликолиз.
При использовании в качестве субстратов гликогена и глюзо-6-фосфата различие между опытом и контролем недостоверно. В то же время, при использовании в качестве субстрата глюкозы, содержание лактата увеличивается в 2—3 раза. Хорошо известно, что при гликолизе в зависимости от субстрата лимитирующим фактором оказываются разные ферменты: при гликогене — фосфорилаза, при глюкозо-6-фосфате — фосфоруктокиназа, при глюкозе — гексокиназа.
Активирование гликолиза только при использовании в качестве субстрата глюкозы говорит о том, что мумие и пентоксил активируют гексокиназу.
Наибольший интерес представляют результаты изучения дыхания митохондрий. Опыты проведены двумя сериями: в марте-апреле, а затем в январе, когда животные наиболее устойчивы.
Митохондрии — клеточные органеллы, «ответственные» за дыхание» клеток, выделялись из гомогенатов печени методом дифференциального центрифугирования. Дыхание изучали полярографически с применением закрытого платинового электрода типа Кларка, используя в качестве субстратов сукцинат и кетоглутарат на сахарозной и солевой средах. Из-за чрезвычайной громоздкости мы не приводим табличное данные, а лишь обсудим результаты исследования. (Желающие могут познакомиться с подробностями в журнале "Вопросы медицинской химии», 1978, 1).
Мумие в дозе 100 мг/кг вызывало достоверное увеличение дыхательной активности на сукцинате в сахарозной среде во всех метаболических состояниях. Наибольшее увеличение активности потребления кислорода наблюдалось при фосфорилировании добавленной АДФ. Соотношение скоростей дыхания в отдыхе и покое увеличивалось в полтора раза. Увеличение скорости дыхания после фосфорилирования, вероятно, свидетельствует об увеличении АТФ-азных реакций расхода синтезирующейся АТФ ввиду усиленного под воздействием мумие биосинтеза.
Процесс потребления кислорода при воздействии разобщителя ДНФ увеличивается не всегда достоверно. На солевой среде наблюдались аналогичные изменения дыхания, однако, в процентном отношении они были меньшими, так как, вероятно, под влиянием самого мумие митохондрии набухают в костном мозге и в селезенке значительно увеличивается количество ретикулярных и плазматических клеток. У доноров же, принимающих в процессе антистафилококковой иммунизации мумие, титры антиальфатоксина нарастали в 2,5—3 раза.
Содержание РНК в печени животных, получавших мумие, достоверно увеличивается, а содержание ДНК не изменяется. Эти данные подтверждаются результатами гистофункционального исследования— усилением пиронинофилии гепатоцитов — и, очевидно, связаны с содержанием нуклеаз, так как накопление нуклеотидов в кроветворных органах крыс, получавших мумие, по данным С. И. Паук и Т. М. Тухтаеаа (1973) происходило параллельно снижение нуклеазной активности.
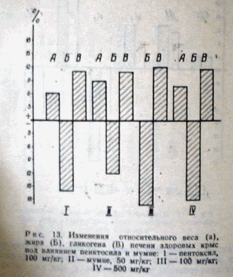
Рис. 13. Изменения относительного веса (а), жира (Б), гликогена (В) печени здоровых крыс под влиянием пентоксила и мумие: I — пентоксил, 100 мг/кг;II — мумие, 50 мг/кг; III — 100 мг/кг; IV — 500 мг/кг
Под влиянием мумие и пентоксила в печени крыс на 12-20% увеличивается содержание гликогена и на 16—20 % снижается содержание жира (рис. 13), что свидетельствует о превышении липолиза над липогенезом и также подтверждается гистофункциональными исследованиями.
Мумие и пентоксил активно изменяют гликолиз (табл. 18).
При использовании в качестве субстратов гликогена и глюзо-6-фосфата различие между опытом и контролем недостоверно. В то же время, при использовании в качестве субстрата глюкозы, содержание лактата увеличивается в 2—3 раза. Хорошо известно, что при гликолизе в зависимости от субстрата лимитирующим фактором оказываются разные ферменты: при гликогене — фосфорилаза, при глюкозо-6-фосфате — фосфоруктокиназа, при глюкозе — гексокиназа.
Активирование гликолиза только при использовании в качестве субстрата глюкозы говорит о том, что мумие и пентоксил активируют гексокиназу.
Наибольший интерес представляют результаты изучения дыхания митохондрий. Опыты проведены двумя сериями: в марте-апреле, а затем в январе, когда животные наиболее устойчивы.
Митохондрии — клеточные органеллы, «ответственные» за дыхание» клеток, выделялись из гомогенатов печени методом дифференциального центрифугирования. Дыхание изучали полярографически с применением закрытого платинового электрода типа Кларка, используя в качестве субстратов сукцинат и кетоглутарат на сахарозной и солевой средах. Из-за чрезвычайной громоздкости мы не приводим табличное данные, а лишь обсудим результаты исследования. (Желающие могут познакомиться с подробностями в журнале "Вопросы медицинской химии», 1978, 1).
Мумие в дозе 100 мг/кг вызывало достоверное увеличение дыхательной активности на сукцинате в сахарозной среде во всех метаболических состояниях. Наибольшее увеличение активности потребления кислорода наблюдалось при фосфорилировании добавленной АДФ. Соотношение скоростей дыхания в отдыхе и покое увеличивалось в полтора раза. Увеличение скорости дыхания после фосфорилирования, вероятно, свидетельствует об увеличении АТФ-азных реакций расхода синтезирующейся АТФ ввиду усиленного под воздействием мумие биосинтеза.
Процесс потребления кислорода при воздействии разобщителя ДНФ увеличивается не всегда достоверно. На солевой среде наблюдались аналогичные изменения дыхания, однако, в процентном отношении они были меньшими, так как, вероятно, под влиянием самого мумие митохондрии набухают— увеличивается их проницаемость — и поэтому разница между контролем и опытом в соленой среде меньше чем в сахарозной. Энергетический процесс в митохондриях под воздействием мумие находится и состоянии активности и солевая среда, как известно, требует дополнительного расхода энергии.
Это предположение подтверждается данными опытов с дозой мумие 500 мг/кг. Скорости дыхания отдыха и покоя не изменяются, а скорость дыхания при фосфорилировании даже снижается, По-видимому, функциональное перенапряжение митохондрий приводит к ингибированию процессов фосфорилирования. Совершенно аналогичные результаты получены у животных, которым вводили пентоксил в дозе
200 мг/кг - у них также отмечалось снижение скорости дыхания при фосфорилировании.
На альфа-кетоглутярате при дозе мумие 100 мг/кг процесс дыхания качественно аналогичен происходящему на сукцинате; мумие в дозе 500 мг/кг и пентоксил в дозе 200 мг/кг на сахарозной среде дают ингибирующий эффект, а на солевой среде этот эффект еще более выражен.
Пентоксил (100 мг/кг) увеличивает дыхательную активность, в меньшей мере, чем мумие и той же дозе. Кроме того, он дает неблагоприятный эффект на НАД-зависимых субстратах — снижается дыхательный контроль и коэффициент фосфорилирования ДНФ.
Нельзя не подчеркнуть, что увеличение дыхательной активности под влиянием дозы мумие 100 мг/кг происходило во всех метаболических состояниях, в двух средах инкубации и но двух различных субстратах и, следовательно, является общей закономерностью.
Описанный биохимические исследования, как уже было указано, полностью коррелируют с данными гистофункционального изучения препаратов печени.
У интактных животных под влиянием пентоксила и мумие нарастало количество купферовских клеток. После приема пентоксила - 30,8±2,74, после 50 мг/кг мумие - 30,4±2,31, а после 500 мг/кг мумие 38,4±2,72 (контроль - 24,7±2,15). Одновременно увеличивались размеры клеточных тел, повышались в них ШИК-реакция и активность кислой фосфатазы. Клетки располагались на некоторых участах цепочками, их ядра становились овальными или даже округлыми. У животных, получавших мумие, независимо от дозы, отмечалось умеренное полнокровие сосудов печени с повышением в стенках активности щелочной фосфазы. По ходу портальных трактов нарастало количество тучных клеток. У животных, получавших пентоксил, описанные изменения были менее выражены.
Структура гепатоцитов при всех видах стимулчции практически не изменялась, но в них отмечалось усиление пирокинофилии. Глыбки РНК были более крупными и ярко окрашенными. Несколько нарастала активность моноаминооксидазы, особенно у животных, получавших пентоксил. Содержание жира в печени несколько снижалось, содержание гликогена чаще было повышено.
У контрольных крыс второй серии через сутки после резекции печени ее относительный вес превосходил исходный на 20%. Учитывая резкое полнокровие органа можно думать, что это превышение веса связано именно с ним. В гепатоцитах при гистоморфологическом исследовании обнаруживались накапливающиеся мелкие и средней величины капли жира между которыми располагались довольно крупные и ярко окрашенные пиронином глыбки РНК. Усиливалась активность кислой и щелочной фосфаз. Активность неспецифичной эстеразы была либо умеренно снижена, либо не изменилась. По ходу портальных трактов отмечалось повышение числа лимфо-гистиоцитарных клеток. Количество купферовских клеток повышалось умеренно, без выраженных метаболических сдвигов в них.
У жиаотных, получавших мумие, через сутки после операции относительный вес печени был равен исходному. Полнокровия не было. Отмечалась несколько большая инфильтрация по ходу портальных трактов и реже в центре долек. Содержание жира в гепатоцитах было ниже, чем в контоле, активность неспецифической эстеразы снижалась. Активность кислой фосфатазы в купферовских клетках была выше, чем в контроле, а щелочной фосфатазы - ниже. Содержание РНК в клетках было значительно выше, чем в контроле, и особенно повышалось на 4 сутки после резекции печени, причем к этому сроку значительно уменьшалось полнокровие и величина инфильтратов, в которых преобладали плазматические клетки.
У животных, получавших пентоксил, относительный вес печени через сутки после резекции также был равен исходному; инфильтрация почти отсутствовала. "Ожирение" гепатоцитов было более выражено, чем у животных, получавших мумие. Активность щелочной фосфатазы и гипертрофия купферовских ж клеток была несколько ниже.
К четвертым суткам инфильтрация нарастала и была больше, чем у контрольных животных, получивших мумие. В элементах инфильтратов активность кислой фосфатазы особенно высока вокруг инфильтратов. Гиперплазия купферовских клеток нарастала с повышением в них ШИК-реакции. Содержание жира оставалось высоким.
Снижение общего белка сыворотки крови через сутки после резекции у контрольных крыс было значительным, а на 4-е сутки почти выравнивалось. Повышались b- и y-глобулины при снижении альбуминов. Белок печени снижался за счет снижения общего и нарастания остаточного азота. Повышался в 1,5 раза коэффициент протеолиза. Через 4 суток все эти изменения оставались значительными.
У крыс, леченных мумие, снижение сывороточного белка было несколько больше, чем в контроле, но белок печени оставался в пределах нормы так же, как и коэффициент протеолиза. У животных, леченных пентоксилом, общий белок сыворотки снижался меньше, чем в контроле, а изменения белка печени и коэффициента протеолиза были даже несколько большими.
В третьей серии опытов крысам проводили 10-дневную затравку НАК. Последняя вызывает так называемую гистотоксическую гипоксию, так как в организме при ее расщеплении высвобождается С-группа. В дальнейшем она связывается с серой, что приводит к нарушениям обмена веществ в результате дефицита сульфгидрильных групп. По данным Л.И. Соловей (1973) затравка НАК вызывает резкое нарушение углеводородного и белкового обмена в печени и проводит к ее жировой дистрофии. (В контрольной группе животных мы получили аналогичные результаты).
При лечении пентоксилом и мумие изменение со стороны протеинограммы, содержания жира и гликогена, содержания жира и гликогена печени были достоверно менее выражены, чем в контроле. Морфологические изменения соответствовали начальной фазе токсического гепатита.
Инфильтрация по ходу портальных трактов у животных, получавших пентоксил, также была выражена меньше, чем в контроле, а в инфильтратах наряду с лимфоидными и гистиоцитарными элементами было много плазматических и тучных клеток и совсем отсутствовали гранулоциты. Гипертрофия купферовских клеток была выражена сильнее, а снижение гликогена было меньше, чем у нелеченных животных.
Эти данные согласуются с сообщениями Р.А. Колесникова (1960), А.Г. Алданозарова с соавторами (1961) о защитном действии пентоксила при свинцовой интоксикации и с данными Р.И. Лифшица (1965, 1966), Н.В. Лазарева (1961), И.Ф. Грех (1970) и других авторов об антиоксидантном и антикатаболическом действии пентоксила.
При лечении мумие инфильтрация также была выражена меньше, чем в контроле, а в инфильтратах обнаружилось много плазматических и тучных клеток и не было гранулоцитов; снижение гликогена было незначительным, а гипертрофия купферовских клеток была выражена еще сильнее, чем при лечении пентоксилом.
Необходимо специально остановиться на результатах определения уровня 11 оксикортикостероидов плазмы у интактных крыс, получавших НБС.
Определение 11-ОКС проводилось флуорометрическим методом (практически у крыс 11-ОКС представлены кортикостероном) по Зильберу-Потеру в модификации Юдаева -Попкова. Уровень гормона у контрольных животных составил 34,3±0,65 мкг%. После приема пентоксила он снизился на 21,8%, после приема мумие в дозе 50 мг/кг — на 34%, а при дозе 500 мг/кг - на 33,8%, то есть, соответственно, до 24,7±0,78, 22,6 ±1,14 и 22,7 ± 1,26 мкг%.
Этот факт — снижение уровня кортикостерона --объясняет многие из обнаруженных эффектов, так как известно, что кортикостерон ингибирует активность гексокиназ, клеточное деление и т. д.
Известно, что в организме животного органические соединения претерпевают ряд метаболических превращений и в результате переходят в неактивную форму. Этот процесс называется инактивацией ксенобиотиков, то есть нейтрализацией вредных для организма веществ. Одним из главных ферментов и этом процессе является цитохром Р-450 печени (А. И. Арчаков, 1974)главный фермент системы микросомального окисления.
С целью определения механизмов детоксицирующего действия мумие; мы изучили его действие на систему микросомального окисления, в частности на содержание в печени ферментов Р-450 и в-5.
Опыты были проведены на 18 белых крысах-самцах весом 120-160 г. На протяжении 10 дней в желудок животным вводили 4% раствор мумие из расчета 100 мг/кг. Череа 48 часов после последнего введения крыс забивали и определяли в печени содержание цитохромов по методу Омура и Сато. Контролем, как обычно, служили животные, которым в желудок вводился физиологический раствор в том же объеме.
Содержание цитохрома Р-450 у контрольных животных составило 0,9:±0,02 нмоль/мг белка микросом, а цитохрома в 5 — 0,89±0,02. У животных, получавших мумие, содержание цитохромов увеличивалось до 2,11 ±0,4 и 2,0±0,4 нмоль/мг белка микросом, соответственно. Таким образом, было доказано стимулирующее действие мумие на систему микросомального окисления печени животных. Этот вывод был подтвержден еще одной серией опытов (рис. 14).
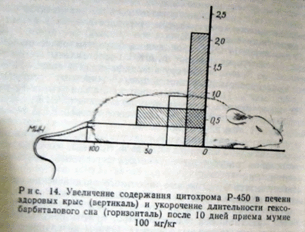
Рис. 14. Увеличение содержания цитохрома Р-450 в печени здоровых крыс (вертикаль) и укорочение длительности гексобарбиталового сна (горизонталь) после 10 дней приема мумие 100 мг/кг
Данные литературы свидетельствуют, что связывание и метаболизм барбитуровых наркотиков происходит исключительно на цитохроме Р-450. В связи с этим 16 крысам внутрибрюшинно вводили гексобарбитал 90 мг/кг в 3% растворе, который готовили непосредственно перед введением. Чеpeз 3—5 минут животные засыпали. Длительность сна регистрировалась с момента перехода животного в положение на боку и составила 109±13,6 минуты. Затем крысам на протяжении 10 дней через зонд в желудок вводили мумие по
100 мг/кг и через сутки после последнего введения вновь определяли у них длительность сна при той же дозе гексобарбитала. Контролем служили данные, полученные у животных, находившихся в тех же условиях, но получавших вместо мумие физиологический раствор. Длительность сна у контрольных животных практически не изменилась (106± 11,3 мин), в то время как у животных, получавших мумие, длительность сна сократилась до 62±12 минут, что полностью коррелирует с увеличением под влиянием мумие цитохрома Р-450.
Таким образом, проведенные нами исследования позволили установить, что саянское мумие обладает анаболическим и антикатаболическим действием не меньшим, чем пентоксил. Энергетическое обеспечение этого эффекта происходит за счет увеличения гликолиза, и, главное, — за счет усиления процессов дыхания и фосфорилирования, преобладания липолиза над липогенезом, а также, вероятно, за счет снижения уровня кортикостерона.
О возросшей активности печени свидетельствуют не только увеличение в ней РНК, но и нарастание количества тучных клеток, которые по мнению В. В. Виноградова и Н. Ф. Воробьевой (1972) являются показателем функциональной активности органа и уровня метаболизма в нем.
При травматическом и токсическом поражении печени пентоксил и в равной мере мумие оказывают защитное действие и стимулируют процессы регенерации.
Действие мумие во многом схоже с действием пентоксила, однако мумие в большей мере стимулирует нуклеиновый обмен и ретикулоэндотелиальную систему печени.
Кроме того, распространенное мнение об абсолютной безвредности НБС, явно не обосновано. При повышении дозы мумие непосредственные эффекты активации обменных процессов еще сохраняются (в большинстве опытов при дозе мумие 500 мг/кг у крыс уже отмечается некоторое снижение показателей), но процессы клеточного дыхания начинают нарушаться. Возможно, это ингибирующее действие больших доз мумие связано с накоплением в ткани печени микроэлементов (рис. 15).

У 20 крыс определялось содержание в ткани печени микроэлементов после 10-дневного приема мумие в дозах 50 и 500 мг/кг. Еще 10 животных, как обычно, получали физиологический раствор. Кроме того, микроэлементный состав был определен в том образце мумие, который был использован в опыте.
Схожесть биологических эффектов, возникающих под влиянием бактериального полисахарида продигиозана, пиримидиновых и пуриновых производных пентоксила и метацила, дрожжевой нуклеиновой кислоты и нуклеината натрия и сложного микроэлементно-органического комплекса, которым является мумие, позволяет думать, что все эти столь различные по своему «химическому строению» препараты, являются своеобразными ключами к системе гомеостаза. (Кстати, с эффектами неспецифических биогенных стимуляторов во многом схожи и эффекты лазерного излучения, интерес к которому «утешил» меня после прощания с мумие).
Эти «ключи» открывают «каналы» в системе саногенеза, нарушенной в процессе заболевания подсистеме гомеостаза в организме, и повышают его возможности справиться с заболеванием — санировать самого себя.
(499) 409-0-418
(905) 771-61-57
Copyright (c) 2010 Mumio.su Все права защищены.
Копирование материалов с данного сайта запрещено и преследуется по закону в соответствии с действующим законодательством РФ